ノルアドレナリンとアドレナリン受容体 ~交感神経を中心に~
アドレナリンとエピネフリン
カテコールアミンと呼ばれる種類の化学物質で分子式はC8H11NO3です。分子構造も非常によく似たアドレナリンは同じカテコールアミンですが、分子式C9H13NO3で、別物です。ノルエピネフリンとノルアドレナリンは、まったくの同一物質についている別名です。同様に、エピネフリンとアドレナリンも同一物質をさす別名です。
関係する神経
交感神経の節後線維終末と支配する器官の間での情報伝達を行う神経伝達物質としてノルアドレナリンが利用されています。また、副腎髄質(ふくじんずいしつ)を支配する交感神経の緊張により、ホルモンとしてアドレナリン(人ではノルアドレナリンではなくアドレナリンが多い)が血中に放出されます。
関係する化学物質
- カテコールアミン(Catecholamine,カテコラミン)
- カテコール核と呼ばれるベンゼン環に水酸基が2つ付いた物にアミノ基がついた構造をしている化学物質類の総称です。ドーパミン、ノルアドレナリン、アドレナリン等が生体で重要な糧コールアミンです。
- ノルアドレナリン(Noradrenalin :Norad)
-
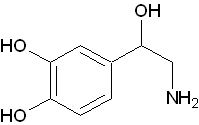 交感神経末梢で放出される神経伝達物質。
交感神経末梢で放出される神経伝達物質。 - アドレナリン(Adrenaline :Adr)
-
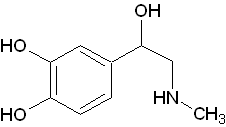 ノルアドレナリン(Norad)が交感神経の神経伝達物質として使われている一方、アドレナリンは副腎髄質から交感神経刺激によって放出されます。
ノルアドレナリン(Norad)が交感神経の神経伝達物質として使われている一方、アドレナリンは副腎髄質から交感神経刺激によって放出されます。 - ドーパミン(Dopamine)
-
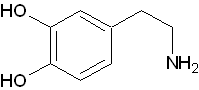 中枢神経系で伝達物質として使われています。末梢神経系では、ノルアドレナリンの生成の前駆体として存在しています。
中枢神経系で伝達物質として使われています。末梢神経系では、ノルアドレナリンの生成の前駆体として存在しています。 - イソプロテレノール(Isoproterenol :IPR)
-
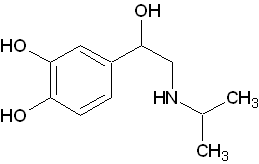 イソプロテレノールは体内で生産される物質ではなく、人工的に合成されたカテコールアミンです。イソプレナリンとも呼びます。アドレナリン受容体であるβ受容体に強く作用します。
イソプロテレノールは体内で生産される物質ではなく、人工的に合成されたカテコールアミンです。イソプレナリンとも呼びます。アドレナリン受容体であるβ受容体に強く作用します。 - アミントランスポーター
- ノルアドレナリンを放出した交感神経末梢のシナプス前線維膜にあるタンパク質の構造物で、細胞外にあるノルアドレナリンを細胞内に取り込みます。放出したノルアドレナリンを回収・再利用するのに役立っています。
- カテコール-O-メチルトランスフェラーゼ(catechol-o-methyltransferase :COMT)
- 細胞外にあって、細胞外に放出されたカテコールアミンを、そのカテコール核の水酸基をメチル化することで不活化する酵素です。COMTのおかげで放出されたカテコールアミンの効果がずっと続くことなく消失します。
- モノアミンオキシターゼ(monoamine oxydase :MAO)
- 細胞内のミトコンドリア中にあり、モノアミントランスポーターによって細胞内に取り込まれたカテコールアミンを不活化します。MAOは、シナプス小胞内のカテコールアミンは不活化できないので、シナプス小胞に取り込まれる前の細胞質中にあるカテコールアミンを不活化します。
交感神経末端(シナプス前部)でのノルアドレナリンの生産・放出
ノルアドレナリンは、チロシン -> ドーパ -> ドパミン -> ノルアドレナリン の順に合成されます。ノルアドレナリンはシナプス前でシナプス小胞に貯蔵されていますが、交感神経刺激が伝導して来ると、細胞外に放出され、シナプス後のアドレナリン受容体(アドレナリンレセプター)に到達します。
ノルアドレナリンは拡散して受容体に到達するので、あまったノルアドレナリンは、細胞外にあるカテコール-O-メチルトランスフェラーゼ(COMT)によってメチル化による不活化がされたり、シナプス前膜上にあるアミントランスポーターによって回収・再利用されます。
細胞内に回収されたノルアドレナリンは、そのまま細胞質中にあると、モノアミンオキシターゼ(MAO)によって分解されてしまいますが、小胞モノアミン・トランスポーターによってシナプス小胞内に取り込まれることによって、分解されずに再利用されます。
アドレナリン受容体 アドレナリンレセプター
アドレナリンレセプターは、臓器、器官の細胞表面にあるタンパク質の構造物で、交感神経終末から放出されたノルアドレナリンを受け取る口です。アドレナリン(Adr)、ノルアドレナリン(Norad)、イソプロテレノール(IPR)等、各種かてこーるあみんに対する反応の違いで、アドレナリン受容体は、α(アルファ)受容体、β(ベータ)受容体と分類され、α受容体はさらにα1受容体、α2受容体、β受容体は、β1受容体、β2受容体、β3受容体と分類されています。ここでは、自律神経と血液循環の観点から押さえておきたい受容体について解説します。
交感神経の心臓支配 β1受容体(ベータ1レセプター)
心臓の拍動数を制御している洞房結節(どうぼうけっせつ)には、交感神経の末梢が多く到達しており、そこではβ1受容体が分布しています。心臓の収縮力をになう心筋(心室筋(しんしつきん)や心房筋(しんぼうきん))にもβ1受容体が分布しています。
β1レセプターはGTP結合タンパク質と結合した受容体で、ノルアドレナリンと結合すると、それに続いて起こる細胞内情報伝達機構を経てカルシウムイオンチャネル(Ca2+)を開きます。すると、Ca2+が細胞内に流入してくるので、細胞を興奮(脱分極)へ傾かせるため、洞房結節は興奮しやすくなり拍動が頻繁になります。
β1レセプターは詳細に研究されていて、反応過程がもっと詳しく分かっています。ここではもう少し詳しく解説します。β1レセプタはアドレナリン等のアゴニストによって活性化すると、結合しているG蛋白によって細胞膜にあるアデニル酸シクラーゼ(Adenylate cyclase)を活性化させ、cAMP(cyclic AMP)を合成させます。このcAMPはプロテインキナーゼA(protein kinase A)を活性化させます。プロテインキナーゼAは、細胞膜上のカルシウムイオンチャネルを活性化(カルシウムの流入を促進させる)させたり、筋小胞体とよばれる細胞内でカルシウムイオンを溜めておく貯蔵庫のカルシウムポンプを活性化させて筋小胞体へのカルシウム貯蓄機能を促進させます。カルシウムイオンは筋肉が収縮する際に直接必要になるものであり、この一連のカルシウム取り込みにより収縮力が増加します。
交感神経の血管平滑筋支配 α1受容体(アルファ1レセプター)、β2受容体(ベータ2レセプター)
血管の平滑筋は血管の収縮力を制御している筋肉です。血管平滑筋が収縮すると、血管は細く固くなるため、血流は細くなり、血圧は上昇に傾きます。反対に、平滑筋がゆるむと、血管は広がり柔らかくなるため、血流は増え、血圧は低下に傾きます。
血管の平滑筋には、部位によりα1受容体とβ2受容体が存在しています。α1受容体もβ2受容体も、GTP結合タンパク質と結合た受容体ですが、α1受容体は、ノルアドレナリンと結合すると、それに続いて起こる細胞内情報伝達機構により、細胞内の筋小胞体からカルシウムイオン(Ca2+)を放出させ、さらに細胞膜にある電位依存型カルシウムチャネルを開くことで細胞外からもカルシウムイオンを流入させます。カルシウムイオンの増加は平滑筋を収縮させます。一方のβ2受容体は、ルアドレナリンと結合すると続いて起こる細胞内情報伝達機構により、これらの作用を阻害するように働きます。つまり、β2受容体は血管平滑筋を弛緩させ(ゆるませ)ます。
補足ですが、β2受容体は、腸などの消化器の平滑筋にも存在しており、交感神経の緊張により平滑筋を抑制させ、消化吸収活動を低下させます。
交感神経活動のネガティブフィードバック α2受容体(アルファ2レセプター)
α2受容体は、交感神経の終末や副交感神経の終末、つまり、シナプス前に存在しています。アルファ2受容体は、GTP結合タンパク質と結合た受容体で、ノルアドレナリンと結合すると、それに続いて起こる細胞内情報伝達機構により、カルシウムイオンチャンネル(Ca2+)が開きにくくなるように働きます。その結果、細胞が脱分極しにくくなり細胞活動は抑制されます。
つまり、交感神経の末端から放出されたノルアドレナリンを受け取ることで、自分自身のノルアドレナリンの放出を抑える(ネガティブフィートバック)働きをします。その一方、副交感神経の末梢にもα2受容体があるために、ノルアドレナリンの放出が多くなると、アセチルコリンの放出が抑えられることになり、こちらは、正のフィードバックがかかることになります。(交感神経末梢のシナプス前には、ムスカリン受容体(M2受容体)があり、アセチルコリンと結合することで細胞を過分極に導き、ノルアドレナリンの放出を抑える効果があります。)
カテコールアミンと各アドレナリンレセプターの作用強度・まとめ
アドレナリン、ノルアドレナリン、イソプロテレノール、ドーパミン等のカテコールアミンは、アドレナリンレセプターと概ね結合し作用しますが、その作用の強弱に差があります。
| レセプタータイプ | α1レセプター | α2レセプター | β1レセプター | β2レセプター |
| 支配器への効果 | Ca2+増加促進 | 脱分極を抑制 | 興奮(脱分極) | Ca2+増加抑制 |
| 部位 | 血管平滑筋 | 自律神経前シナプス | 心臓 | 気管・腸・血管平滑筋 |
| ノルアドレナリン作用 | 強 | 強 | 強 | 弱 |
| アドレナリン作用 | 強 | 強 | 強 | 強 |
| イソプロテレノール作用 | 作用しない | 作用しない | 強 | 強 |